Глава 36 МЕХАНИЗМ ЗРЕНИЯ
Глава 36
МЕХАНИЗМ ЗРЕНИЯ
§ 1. Ощущение цвета
§ 2. Физиология зрения
§ 3. Палочки
§ 4. Сложные глаза насекомых
§ 5. Другие типы глаз
§ 6. Нервные механизмы зрения
§ 1. Ощущение цвета
Обсуждая механизм зрения, прежде всего необходимо понять, что мы обычно видим не беспорядочный набор цветных или световых пятен (разумеется, если не находимся на выставке некоторых современных художников!). Когда мы смотрим на что-то, то видим человека или вещь; другими словами, мозг интерпретирует то, что мы видим, как человека или вещь. Как он это делает — никому неведомо, но делает он это, надо сказать, великолепно. Хотя мы на опыте учимся узнавать, как выглядит человек, однако есть некоторые более элементарные свойства зрения, которые тем не менее тоже включают сопоставление информации от различных частей того, что мы видим. Чтобы понять, как происходит интерпретация изображения в целом, следует изучить первые стадии сопоставления информации от различных клеток сетчатки. В настоящей главе мы сконцентрируем наше внимание главным образом именно на этих сторонах зрения, хотя попутно упомянем и о некоторых других смежных вопросах.
Примером такого сопоставления информации (хотя и на элементарном уровне), поступающей одновременно от нескольких частей глаза и происходящей помимо нашей воли, контроля и сознания, может служить голубая тень от белого света, когда одновременно экран освещается еще и красным светом. При этом по меньшей мере предполагается, что нам известно, что основа экрана красная, и хотя в глаз попадают только «белые» лучи, однако где-то эти кусочки информации складываются вместе и мы видим голубую тень.

Фиг. 36.1. При вращении этого диска одно из колец кажется цветным; при изменении направления вращения окрашенным кажется второе кольцо.
Чем полнее и привычнее картина, тем большую поправку делает глаз. Действительно, Ланд показал следующее: если мы возьмем два диапозитива, поместим перед ними два фильтра, поглощающие красный и белый свет в различных отношениях, мы будем смешивать разные интенсивности кажущегося голубого и красного света и сможем получить довольно правдивое изображение реальной сцены с натуральными предметами. Кроме красного и белого, мы в этом случае получим множество промежуточных цветов. Аналогичные результаты можно получить, смешивая красный и зелено-голубой цвета; оказывается, что мы получаем почти полный набор цветов. Впрочем, если внимательно приглядеться к ним, то мы увидим, что они не столь уж хороши. Но даже и при этих условиях просто удивительно, как много можно получить только из красного и белого цвета. Чем больше изображение напоминает реальную картину, тем больше мы способны компенсировать то обстоятельство что цвет-то фактически только розовый!
Другим примером может служить появление «цвета» на черно-белом вращающемся диске, изображенном на фиг. 36.1. При вращении диска смена черного и белого цвета для каждого радиуса в точности одинакова; это составляет фон, на котором видны два «кольца». Первое кольцо кажется окрашенным в один цвет, а второе — в другой. До сих пор никто не понимает причины появления здесь окраски, однако ясно, что наиболее правдоподобное объяснение состоит в том, что на каком-то элементарном уровне, по-видимому, в самом глазе происходит сложение информации.
Почти все современные теории цветового зрения сходятся на том, что опыты по смешиванию цветов указывают на существование в глазе только трех сортов пигментов и что ощущение цвета создают именно спектры поглощения этих трех пигментов. Однако полная чувствительность, связанная с характеристиками поглощения этих пигментов, функционирующих одновременно, не обязательно равна сумме их чувствительностей.
Каждый знает, что желтый цвет не кажется нам красновато-зеленым, и многих, вероятно, несказанно удивит тот факт, что каждый видимый ими цвет есть по сути дела смесь разных цветов, ибо им кажется, что чувство цвета вызывается каким-то другим механизмом, а не просто смешиванием, наподобие сочетания звуков в аккорды в музыке, когда одновременно звучат, скажем, три ноты. Ведь в аккорде, если внимательно прислушаться, можно различить отдельные ноты, а в желтом цвете, сколько бы мы ни приглядывались, увидеть красный и зеленый цвет отдельно невозможно.
Уже первые теории зрения утверждали, что имеются три сорта пигментов и соответственно три сорта колбочек, каждая из которых содержит один пигмент; что от каждой колбочки в мозг идут нервы, так что в мозг переносятся три сорта информации и там что-то происходит. Конечно, это очень несовершенная теория, ибо она не позволяет обнаружить, что за информация переносится по зрительным нервам в мозг; она даже не приступала к решению этой проблемы. Мы должны задать более фундаментальный вопрос: не все ли равно, где происходит сложение информации? Насколько необходимо, чтобы эта информация передавалась по зрительным нервам прямо в мозг, и не может ли какой-то первоначальный анализ выполняться самой сетчаткой? Мы знаем, что сетчатка невероятно сложна и обладает множеством внутренних связей (см. фиг. 35.2); какой-то анализ она способна выполнить.
Дело в том, что ученые, занимающиеся анатомией и развитием глаза, показали, что сетчатка, в сущности, не что иное, как часть самого мозга; при развитии зародыша часть мозга выносится вперед, из нее назад вырастают длинные волокна, которые связывают ее с остальным мозгом. По своей организации сетчатка весьма похожа на мозг. По этому поводу кто-то прекрасно сказал, что это «мозг выдумал, как ему взглянуть на мир». Глаза — это кусочек мозга, которым он, так сказать, «касается света», внешнего мира. Таким образом, нет ничего необычного в том, что какой-то анализ цвета происходит уже в самой сетчатке.
Это предоставляет нам весьма интересную возможность. Ведь никакой другой орган чувств не делает столько, если так можно выразиться, предварительных вычислений, как глаз, прежде чем сигнал попадет в нерв, где его можно измерить. Вычисления для всех остальных органов чувств обычно производит сам мозг, а из-за огромного количества внутренних связей в мозге добраться до этого специфического места и произвести какие-то измерения очень трудно. В глазе вычисления производятся в трех слоях клеток, затем результат вычисления передается по зрительному нерву в мозг. Так что здесь мы, по-видимому, впервые получаем возможность физиологически наблюдать, как работает первый слой мозга, быть может, на начальной стадии. Это вдвойне интересно не только для понимания зрения, но и для всех проблем физиологии.
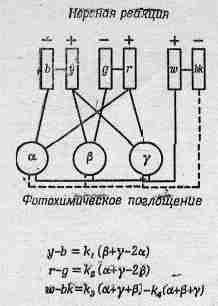
Фиг. 36.2. Нервные связи, согласно теории цветового зрения. b — голубой; y—желтый; g — зеленый; r —красный; w — белый; bk — черный.
Тот факт, что существуют три сорта пигментов, вовсе не означает, что должно быть также три сорта ощущений. Существует теория цветового зрения, основанная на совершенно противоположной цветовой схеме (фиг. 36.2). Согласно этой схеме, какое-то из нервных волокон несет много импульсов, если мы видим желтый цвет, и меньше, чем обычно, если мы видим голубой. Другое нервное волокно точно таким же образом переносит информацию о зеленом и красном цвете, а третье — о белом и черном. Другими словами, в этой теории уже начинают делаться догадки о системе связи и методе анализа.
Вопросы, которые мы пытаемся решить с помощью догадок об этом первоначальном анализе, следующие: проблема кажущихся цветов на розовом фоне; что происходит, когда глаз привыкает к различным цветам; и вопрос о так называемых психологических явлениях. Под этим термином мы понимаем, например, что белый цвет не «ощущается» нами как смесь красного, желтого и синего, и такая теория возникла потому, что, как утверждают психологи, существуют четыре кажущихся чистых цвета: «Существуют четыре мощных возбудителя, вызывающие соответственно простые голубой, желтый, зеленый и красный оттенки. В отличие от таких красок, как сиена, пурпур, фуксин или другие различимые цвета, эти простые оттенки являются несмешанными в том смысле, что ни один из них не принимает участия в образовании других, в частности голубой цвет нельзя назвать желтоватым, красноватым, зеленоватым и т. д.; психологически они представляют первичные оттенки».
В этом состоит так называемый психологический факт. Чтобы выяснить, откуда взялся этот факт, нужно очень старательно просмотреть всю литературу. Все, что мы находим в современной литературе по этому вопросу, повторяет те же утверждения или утверждения одного из немецких психологов, авторитетом которого является Леонардо да Винчи, хорошо всем известный великий художник. Этот психолог говорит: «Леонардо считал, что существует пять цветов». Дальнейшие поиски приводят к еще более древним книгам. В этих книгах говорится примерно следующее: «Фиолетовый цвет — это красновато-голубой, оранжевый — это красновато-желтый, но можно ли красный рассматривать как фиолетово-оранжевый? Не будут ли красный и желтый более основными цветами, чем фиолетовый и оранжевый? На вопрос, какие цвета они считают основными, большинство людей назовут красный, желтый и синий, а некоторые добавят к этим трем еще и четвертый — зеленый. Психологи привыкли принимать эти четыре цвета за основные».
Итак, с точки зрения психологов, раз все говорят, что есть три цвета, а кое-кто утверждает, что четыре, и хотят, чтобы было четыре, ну пусть будет четыре. Это иллюстрирует трудности, сопровождающие психологические исследования. Ясно, что мы таким чувством обладаем, но узнать о нем немного больше очень трудно.
Можно идти по другому пути — физиологическому — и экспериментально выяснить, что на самом деле происходит в мозге, в глазе, в сетчатке или другом каком-то месте, и, может быть, удастся обнаружить, что некоторые комбинации импульсов от различных клеток передаются по определенным нервным волокнам. К сожалению, первичные пигменты не сосредоточены каждый в отдельной клетке: могут быть клетки, в которых содержится смесь различных пигментов, клетки с красным и зеленым пигментами или со всеми тремя сразу (информация об этих трех пигментах будет «белой» информацией) и т. д. Есть много способов связать всю эту систему, и мы должны выяснить, какой из них предпочла природа. В то же время хочется надеяться, что, поняв физиологические связи, мы хоть немного продвинемся вперед в понимании некоторых психологических аспектов. Итак, вперед по этому пути!
§ 2. Физиология зрения
Мы начали говорить не только о цветовом зрении, но о зрении вообще только для того, чтобы напомнить о внутренних связях в сетчатке, показанных на фиг. 35.2. Сетчатка поистине напоминает поверхность мозга. Хотя настоящая картина под микроскопом выглядит несколько более сложно, чем этот схематический рисунок, но тем не менее при тщательном анализе можно увидеть все эти внутренние связи. Дело не в том, что одна часть сетчатки связана с другими частями и что информация, переданная по длинным аксонам, образующим зрительный нерв, представляет собой комбинированную информацию от многих клеток. Дело в том, что существуют три слоя клеток, функции которых следующие: во-первых, это фоторецепторы, на которые непосредственно действует свет, затем промежуточные клетки, которые принимают информацию от одного или нескольких фоторецепторов и снова отдают ее нескольким клеткам третьего слоя, а затем в мозг. Между клетками различных слоев существуют разнообразные перекрещивающиеся связи.
Вернемся к некоторым аспектам строения и функции глаза (см. фиг. 35.1). Свет фокусируется главным образом роговицей, благодаря тому, что поверхность ее искривлена и она «загибает» лучи света. Вот почему под водой мы видим не так хорошо, ибо показатели преломления роговицы (1,37) и воды (1,33) разнятся недостаточно сильно. Позади роговицы находится практически водная среда с показателем преломления 1,33, а дальше — хрусталик, строение которого очень интересно: он состоит из целого ряда слоев, как луковица, с той только разницей, что эти слои прозрачные и показатель преломления их меняется от 1,40 в середине до 1,38 по краям. (Неплохо было бы изготовить линзу с необходимым показателем преломления в любом месте; тогда нам незачем было бы так искривлять ее, как это делается с линзой с постоянным показателем преломления.) Более того, форма роговицы вовсе не сферическая. Сферическая линза обладает известной сферической аберрацией. Наружная часть роговицы более «плоская», чем у сферы, причем как раз настолько, чтобы сферическая аберрация ее оказалась меньше аберрации той сферической линзы, которую мы поставили бы вместо нее! Посредством этой оптической системы роговица — хрусталик свет фокусируется на сетчатку. Если мы смотрим на близко расположенные или удаленные предметы, то хрусталик искривляется или выпрямляется, изменяя тем самым фокусное расстояние и настраиваясь на различную удаленность. Для регулирования общего количества света в глазе имеется радужная оболочка, или радужка, которая определяет «цвет» глаз — у кого карие, у кого голубые. При увеличении количества света оболочка сжимается и зрачок уменьшается, при уменьшении — оболочка расходится и зрачок увеличивается.
Рассмотрим теперь изображенный на фиг. 36.3 нервный механизм, регулирующий аккомодацию хрусталика, движение глаза (способность глаза поворачивать глазное яблоко в глазнице) и диаметр зрачка. Основная часть всей информации попадает в зрительный нерв А, который разделяется на два пучка (о них мы еще будем говорить), и по ним идет в мозг. Однако имеется несколько волокон (именно они сейчас нам и интересны), которые не идут прямо в зрительную кору, где мы «видим» изображение, а вместо этого отправляются в средний мозг Н. Это как раз те волокна, по которым передается информация о средней освещенности и приказы о необходимом диаметре зрачка или, если изображение кажется расплывчатым, о кривизне хрусталика.
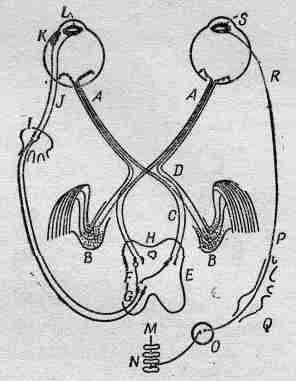
Фиг. 36.3. Нервные связи, управляющие механическими движениями глаза.
Если же изображение раздвоено, то по этим волокнам посылается приказ подрегулировать глаза для бинокулярного зрения. Во всяком случае, они проходят через центр мозга и возвращаются назад в глаз. Буквой K обозначены мышцы, которые управляют хрусталиком при аккомодации, а буквой L — ресничные мышцы. Радужка имеет две мышечные системы: 1) мышца, суживающая зрачок (циркулярная мышца) L; она работает очень быстро и связана непосредственно с мозгом коротким аксоном; 2) мышца, расширяющая зрачок (радиальная мышца), которая действует тогда, когда освещение предмета уменьшается и циркулярная мышца расслабляется. Как и во многих других частях тела, здесь тоже пара мышц работает в противоположных направлениях; почти в каждом таком случае управляющая ими нервная система «настроена» настолько точно, что, когда одной из них посылается приказ сжаться, другой автоматически посылается приказ расслабиться. Однако радужка представляет любопытное исключение: нервы, которые заставляют оболочку сжиматься, мы только что описали, но до сих пор никому точно не известно, откуда выходят нервы, заставляющие ее разжиматься. Они идут куда-то вниз, в спинной мозг в области грудной клетки, из спинного мозга вверх через шейный нервный узел, потом опять назад в голову и к другому концу радужки. Сигнал фактически проходит через совершенно другую нервную систему, не через центральную, а через симпатическую. Очень странно, для чего все это нужно.
В глазе, как мы подчеркивали, имеется еще одна странность: светочувствительные клетки расположены в сетчатке в глубине, так что, прежде чем попасть в рецепторы, свет должен пройти через несколько слоев других клеток: сетчатка как бы вывернута наизнанку! В общем некоторые вещи в устройстве глаза кажутся нам великолепными, а некоторые — просто глупыми.
На фиг. 36.4 показана связь глаза с частью мозга, наиболее непосредственно принимающей участие в процессе зрения.
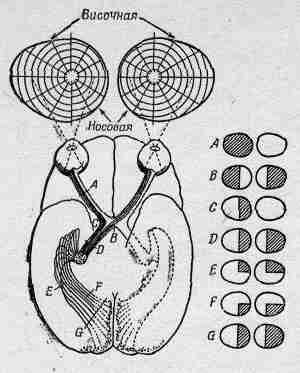
Фиг. 36.4. Нервная связь глаза со зрительной корой.
Зрительные нервные волокна идут в некоторую область, лежащую сразу же за участком D, называемым латеральным коленчатым телом, а затем в участок мозга, называемый зрительной корой. Следует помнить, что от каждого глаза некоторые волокна направляются в другую половину мозга, так что представленная картина не полна. Зрительные нервы от левой части правого глаза проходят через зрительный перекрест B, тогда как нервы от левой части левого глаза обходят его сбоку. Таким образом, левая часть мозга получает всю информацию, идущую от левых сторон обоих глаз, т. е. правой стороны поля зрения, тогда как правая сторона мозга «видит» левую часть поля зрения. Вот каким способом происходит сложение информации от обоих глаз и определяется удаленность предмета. Такова система бинокулярного зрения.
Очень интересны связи между сетчаткой и зрительной корой. Если мы в сетчатке каким-то образом возбудим или разрушим некоторую область, то умирает все волокно, поэтому мы можем узнать, куда оно идет, с чем оно связано. Самое интересное то, что между сетчаткой и зрительной корой, оказывается, существует однозначное соответствие: каждому пятну на сетчатке соответствует пятно в зрительной коре, и два рядом расположенных пятна на сетчатке окажутся рядом и в зрительной коре. Так что зрительная кора, кроме всего прочего, отражает и пространственное расположение палочек и колбочек, хотя и очень искаженно. Предметы, находящиеся в центре поля зрения и занимающие очень мало места на сетчатке, в зрительной коре распространяются на очень много клеток. Ясно, что очень важно, чтобы первоначально расположенные близко предметы оказались так же близко расположенными и в зрительной коре. Однако самое интересное здесь вот что. Участок, который, казалось бы, наиболее важен для близко расположенных предметов, находится как раз в середине поля зрения. Поистине невероятно, но прямая вертикальная линия в середине поля зрения обладает тем свойством, что информация, полученная от всех точек, расположенных справа от нее, поступает в левое полушарие мозга, а информация от точек, расположенных слева,— в правое полушарие. Но прямо посредине проходит граница, так что предметы, которые очень близки и расположены в середине по разные стороны от границы, в мозге оказываются очень далекими! Информация через какие-то другие каналы все же как-то проходит из одной стороны мозга в другую, и это очень странно.
Очень интересно, как все это связывается вместе. Вопрос о том, что уже связано и что еще нужно научиться связывать, довольно стар. Прежде думали, что, по-видимому, никаких врожденных связей вообще нет; имеются только какие-то грубые наметки, и лишь потом на опыте еще в детстве постигают, что когда предмет находится «вон там», то это дает такое-то ощущение. (Врачи постоянно уверенно заявляют о том, что чувствуют маленькие дети, но откуда сами они знают, что чувствует годовалый ребенок?) Может быть, годовалый ребенок, видя предметы «вон там», испытывает какое-то чувство и учится протягивать руку именно «туда», потому что когда он протягивает ее «сюда», то схватить предмет не удается. Но, по-видимому, этот подход все же неверен, ибо, как мы уже видели, во многих случаях такие специфические промежуточные связи существуют уже с рождения.
Более показательны в этом отношении замечательные опыты над саламандрами. (К счастью, у саламандры имеется прямая перекрестная связь без зрительного перекреста, поскольку у нее глаза расположены по бокам головы и поля зрения обоих глаз не перекрываются. Саламандрам поэтому бинокулярное зрение ни к чему.) Опыты эти состоят в следующем. Мы можем перерезать зрительный нерв у саламандры, но он, однако, снова начнет расти из глаз. Так будут восстанавливаться сами собой тысячи и тысячи клеток. И хотя волокна зрительных нервов не будут лежать рядом (они теперь напоминают большой небрежно изготовленный телефонный кабель, все волокна которого перекручены и перепутаны), однако, достигнув мозга, они снова расположатся в надлежащем порядке. Когда перерезают зрительный нерв саламандры, то возникает вопрос: восстанавливается ли он снова? Да, восстанавливается. Таков замечательный ответ. Если саламандре перерезать зрительный нерв, то он снова вырастает и она будет видеть ничуть не хуже, чем раньше. Однако если мы перережем зрительный нерв и перевернем глаз, а потом оставим его в покое, то нервы снова вырастут и саламандра прозреет, но теперь она будет совершать ужасные ошибки: увидев муху вверху, саламандра прыгнет вниз, и она уже никогда не сможет «выучиться» действовать правильно. Так что каким-то непостижимым образом тысячи и тысячи клеток нервных волокон находят в мозге свое истинное место.
Проблема связей в мозге, т. е. в какой степени все там связано и в какой нет,— важнейшая проблема в теории развития живых существ. Ответ еще неизвестен, но его интенсивно ищут.
Аналогичный опыт с золотой рыбкой приводит к тому же результату: в том месте, где мы перережем нерв, образуется страшный узел, подобно большому шраму или опухоли, и, несмотря на все это, волокна снова «прорастут» в мозг к своему истинному месту. Для того чтобы это произошло, волокна, поскольку они растут по старому каналу зрительного нерва, «должны решать», в каком направлении расти. Но как им удается это делать? Возможно, что здесь работает какой-то химический механизм, который по-разному действует на разные волокна. Подумать только, сколь огромно число растущих волокон и каждое из них как-то, по-своему отличается от соседних; реагируя на какой-то химический механизм, оно делает это достаточно однозначно, чтобы отыскать свое истинное место среди окончательных связей в мозге! Это поразительно, фантастично! Это одно из величайших явлений, открытых биологами за последнее время, и оно, несомненно, связано со многими старыми нерешенными проблемами роста, организации и развития организма, особенно зародыша.
Другое интересное явление связано с движением глаза. Чтобы добиться совпадения двух изображений, глаз должен обладать способностью двигаться. Эти движения могут быть разного рода: когда мы следим за чем-то, оба глаза должны поворачиваться одновременно в одном направлении — вправо или влево; когда мы следим за удаляющимся или приближающимся предметом, глаза должны двигаться в противоположных направлениях. Нервы, подходящие к мышцам глаза, как раз приспособлены для этих целей. Одни нервы заставляют наружные мышцы, например левого глаза и внутренние мышцы правого, сокращаться, а противоположные мышцы — расслабляться, так что оба глаза движутся в одну сторону. Но есть и другие центры, возбуждение которых заставляет глаза двигаться навстречу друг другу. Любой глаз может быть скошен в уголок, если второй при этом движется к носу, но совершенно невозможно сознательно или несознательно одновременно повернуть оба глаза в разные стороны, и вовсе не потому, что нет мышц, способных сделать это, а потому, что нет способа послать такие сигналы, чтобы оба глаза отвернулись в разные стороны. (Разумеется, если не произошло никакого нарушения, например не перерезан нерв.) И хотя мышцы одного глаза вполне могут поворачивать его как угодно, даже йоги никаким усилием воли не могут повернуть оба глаза в разные стороны. Просто потому, что нет никакой возможности сделать это. В какой-то мере мы уже скованы от рождения. Это очень важный пункт, ибо большинство прежних книг по анатомии и психологии не признавало или не замечало того факта, что мы в такой степени скованы с самого рождения; они утверждали, что можно всему научиться.
§ 3. Палочки
Посмотрим теперь подробнее, что происходит в палочках сетчатки. На фиг. 36.5 показана микрофотография середины палочки (конец ее выходит вверх за пределы снимка). Справа в увеличенном виде слой за слоем видны плоские структуры, содержащие родопсин (зрительный пурпур) — красящее вещество, или пигмент, который, собственно, и обусловливает функцию палочек. Родопсин представляет собой большие молекулы белка, содержащего специальную группу, называемую ретиненом, которая может быть отщеплена от белка, что, несомненно, и является главной причиной поглощения света. Нам пока не понятно, почему эти структуры плоские, но весьма возможно, что это сделано для того, чтобы молекулы родопсина лежали параллельно друг другу. Химия этого явления известна сейчас довольно хорошо, но, кроме того, возможно, что здесь принимает участие и физика. Может оказаться, что все молекулы располагаются в своего рода ряд, и когда одна из них возбуждается, то вылетевший при этом электрон (или просто волна возбуждения) доходит до некоторого места в конце структуры и порождает сигнал или что-нибудь аналогичное. Это очень важная область, и она еще совсем не разработана. Это поле деятельности для биохимии и физики твердого тела или чего-то в этом духе.
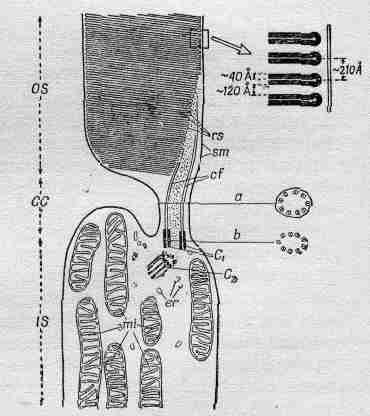
Фиг. 36.5. Электронная микрофотография палочки.
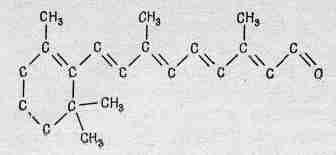
Фиг. 36.6. Строение ретинена.
Те же самые слоистые структуры найдены и в других местах, где тоже важен свет, например в хлоропласте растений, где под действием света происходит фотосинтез. При большом увеличении мы обнаруживаем те же самые слои, но, конечно, вместо ретинена мы находим хлорофилл. Химическая форма ретинена показана на фиг. 36.6. Его боковая ветвь содержит серию альтернирующих двойных связей, характерную почти для всех сильно поглощающих органических веществ, подобных хлорофиллу, гемоглобину и т. д. Эти вещества человек не может изготовить в своих собственных клетках и должен получать их с пищей в виде специального вещества, в точности похожего на ретинен, за исключением водородной связи на правом конце. Называется это вещество витамином А. Если в пище его недостаточно, то запас ретинена в организме не пополняется и развивается то, что мы называем куриной слепотой, т. е. количества пигмента будет недостаточно для того, чтобы можно было видеть в сумерках.
Известно также, почему такая серия двойных связей очень сильно поглощает свет. Я немного расскажу вам об этом. Альтернирующая серия двойных связей называется сопряженной двойной связью. Двойная связь означает, что там есть дополнительный электрон, который легко сдвинуть вправо или влево. Когда свет ударяет по этой молекуле, то электрон каждой двойной связи на один шаг сдвигается. В результате сдвинутся электроны во всей цепи, подобно тому, как упадут при толчке поставленные друг за другом костяшки домино, и хотя каждый из них проходит очень небольшое расстояние (мы считаем, что в отдельном атоме электрон может проходить только очень маленькое расстояние), в целом получается такой же эффект, как будто электрон с одного конца перескочил на другой! Это то же самое, как если бы один электрон прошел все расстояние взад и вперед, а в таком случае происходит значительно более сильное поглощение под действием электрического поля, чем если бы мы передвинули электрон только на расстояние, связанное с одним атомом. А поскольку двигать электрон взад и вперед не так уже трудно, то ретинен очень сильно поглощает свет; таков механизм, в основе которого лежит физика и химия.
§ 4. Сложные глаза насекомых
Вернемся теперь к биологии. Человеческий глаз — отнюдь не единственный тип глаза. Хотя глаза почти всех позвоночных похожи на человеческие, однако у низших животных мы встречаем множество других типов глаз. Обсуждать их у нас нет времени. Но среди беспозвоночных (например, у насекомых) встречаются и высокоразвитые типы глаз; это сложные, или фасеточные, глаза. (У большинства насекомых, кроме больших: фасеточных глаз, есть еще простые глаза, или глазки.) Наиболее тщательно изучалось зрение пчелы. Изучать особенности зрения пчел легко, ибо их, как известно, привлекает мед и мы можем ставить опыты, смазывая медом, например, голубую или красную бумагу и наблюдая, какая из них привлечет пчелу. Этим методом были обнаружены очень интересные особенности зрения пчелы.
Прежде всего, пытаясь определить, насколько отчетливо пчела видит разницу между двумя кусочками «белой» бумаги, некоторые исследователи нашли, что она видит ее не очень хорошо, а другие, наоборот, что она делает это чертовски здорово. Даже если брались два почти в точности одинаковых кусочка бумаги, пчела все же различала их. Один кусок бумаги, например, отбеливался цинковыми белилами, а другой — свинцовыми, и, хотя оба они выглядели в точности одинаково, пчела различала их, ибо они по-разному отражают ультрафиолетовый свет. Таким образом, было обнаружено, что глаз пчелы чувствителен к более коротким длинам волн, чем глаз человека. Наши глаза видят в диапазоне от 7000 до 4000 Е, от красного до фиолетового, а пчелы могут видеть вплоть до 3000 Е, т. е. в ультрафиолетовой области! А это порождает целый ряд очень интересных эффектов. Прежде всего пчелы различают многие цветы, которые нам кажутся абсолютно одинаковыми. В этом нет ничего удивительного; ведь цветы цветут вовсе не для того, чтобы радовать наш взор. Они служат приманкой для пчел, своеобразным сигналом о том, что здесь есть мед. Всем известно, что есть очень много «белых» цветов. Цвет, который нам кажется белым, по-видимому, пчелы не видят, ибо выяснилось, что разные белые цветы не отражают ультрафиолетовые лучи столь полно, как это делают истинно белые цветы. От белого предмета отражается не весь падающий на него свет, ультрафиолетовые лучи теряются, а это в точности то же, что для нас потеря голубого цвета, т. е. получение желтого цвета. Итак, все белые цветы кажутся пчелам цветными. Однако нам известно также, что пчелы не видят красного цвета. Выходит, можно считать, что красные цветы кажутся пчелам черными? Ничего подобного! Тщательное изучение красных цветов показывает, что, во-первых, даже наши глаза способны различать у огромного большинства красных цветов легкий синеватый оттенок, вызванный дополнительным отражением большинством из них синего цвета, который находится в видимой пчелами области. Кроме того, опыты также показывают, что цветы отличаются по своей способности отражать ультрафиолет от различных частей лепестка и т. д. Итак, если бы мы могли видеть цветы такими, как их видят пчелы, то нашли бы их еще прекраснее и разнообразнее!
Впрочем, было обнаружено, что имеются такие красные цветы, которые не отражают голубых или ультрафиолетовых лучей, поэтому они должны казаться пчелам черными! Это в какой-то степени объясняет недоумение тех людей, которых очень волнует этот вопрос: ведь черный цвет не кажется привлекательным, да и трудно его отличить от грязной густой тени. Так оно и получается на самом деле: пчелы не прилетают на эти цветы. Но зато как раз они нравятся крохотным колибри; оказывается, эти птички отлично видят красный цвет!
Еще одна интересная сторона зрения пчелы. Взглянув на кусочек голубого неба и не видя самого солнца, пчела, по-видимому, может все-таки определить, где находится солнце. Для нас это не так-то просто. Посмотрите из окна на небо. Вы видите, что оно голубое. А в каком направлении находится сейчас солнце? Пчела может это определить, ибо она очень чувствительна к направлению поляризованного света, а отраженный от неба свет поляризован. До сих пор спорят о том, как это ей удается: то ли потому, что рассеяние света различно при разных обстоятельствах, то ли глаза пчелы непосредственно чувствительны к направлению поляризованного света. В самое недавнее время получены данные о непосредственной чувствительности глаза пчелы.
Говорят также, что пчела способна различать отдельные вспышки света с частотой 200 раз в 1 сев, тогда как мы различаем только 20 вспышек. В улье пчелы движутся очень быстро; они шевелят лапками, машут крылышками, но наши глаза с трудом успевают уследить за всеми этими движениями. Вот если бы мы могли различать более быстрые мелькания, тогда другое дело. Видимо, для пчелы очень важно, чтобы ее глаза имели столь быструю реакцию.
Теперь поговорим о том, какова, собственно, острота зрения у пчелы? Глаз пчелы сложный; состоит он из огромного числа особых глазков, называемых омматидиями, которые расположены на почти сферической поверхности по бокам головы насекомого.
На фиг. 36.7 показан омматидий. В его вершине находится прозрачная область, своего рода «хрусталик», но в действительности это больше напоминает фильтр, заставляющий свет идти вдоль узкого волокна, где, по-видимому, и происходит его поглощение. От другого его конца отходит нервное волокно. Центральное нервное волокно имеет по бокам шесть клеток, от которых по сути дела оно и отходит. Для наших целей этого описания вполне достаточно; главное то, что клетка имеет коническую форму, и множество таких клеток, примыкая друг к другу, образует поверхность глаза пчелы.
Посмотрим теперь, каково разрешение такого глаза. Проведем линию (фиг. 36.8), схематически представляющую омматидий, на поверхности глаза, который мы будем считать сферой радиусом r. Мы сейчас попытаемся вычислить ширину каждого омматидия, для чего напряжем немного нашу сообразительность и предположим, что природа столь же сообразительна, как и мы! Если омматидий очень велик, то разрешение не может быть большим. Иначе говоря, один омматидий получает информацию об одном направлении, соседний — о другом и т. д., а предметы, попавшие в промежуток, пчела не сможет видеть достаточно хорошо. Таким образом, неопределенность остроты зрения глаза, несомненно, связана с угловым размером конца омматидия относительно центра кривизны глаза. (На самом деле глаза расположены только на поверхности головы.) Но угол от одного омматидия до следующего равен, конечно, диаметру омматидия, деленному на радиус кривизны поверхности глаза:
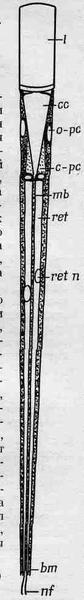
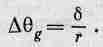
Фиг. 36.7. Строение омматидия.
(36.1)
Итак, можно сказать: «Чем меньше будет значение d, тем больше окажется острота зрения.
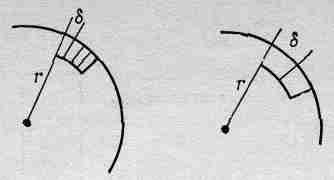
Фиг. 36.8. Схема распределения омматидиев на поверхности глаза пчелы.
Но почему же тогда природа не дала пчеле очень-очень мелкие омматидии?» В ответ можно сказать следующее: мы уже достаточно хорошо знаем физику, чтобы понять, что при попытке пропустить свет через узкую щелку из-за дифракции невозможно достаточно хорошо видеть в данном направлении, ибо туда будет попадать свет от различных направлений, т. е. от всех направлений, находящихся внутри угла Dqd, такого, что
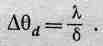
(36.2)
Теперь ясно, что, если взять б слишком маленьким, каждый омматидий вследствие дифракции будет видеть не только в одном направлении! Но если сделать значение d слишком большим, то, хотя каждый будет смотреть в одном направлении, их окажется слишком мало, чтобы получить достаточно подробную картину. Таким образом, мы должны подобрать такое расстояние d, чтобы полный эффект этих двух механизмов был минимальным. Если мы сложим два выражения и найдем место, где сумма имеет минимум, то получим
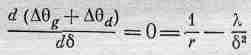
(36.3)
что дает расстояние
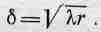
(36.4)
Если мы для оценки будем считать, что r приблизительно равно 3 мм, а длину волны света, который пчела видит, примем равной 4000 А, то после умножения и извлечения корня найдем
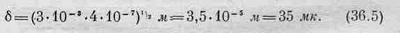
В книгах указывается диаметр, равный 30 мк. Как видите, согласие оказывается довольно хорошим! Ясно, что именно этот механизм определяет размер глаза пчелы и он вполне доступен нашему пониманию. Подставив теперь полученное число в (36.1), легко определить, каково угловое разрешение глаза пчелы. Оно оказывается очень плохим по сравнению с глазом человека. Мы способны видеть вещи, кажущийся размер которых в тридцать раз меньше, чем видит пчела. Так что по сравнению с человеком изображение у пчелы получается довольно расплывчатое, несфокусированное.
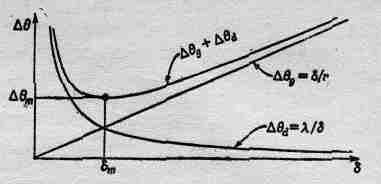
Фиг. 36.9. Оптимальный размер омматидия, равный dm.
Тем не менее так оно есть, и на большее она просто не может рассчитывать. Естественно возникает вопрос: а почему бы пчеле не завести себе такой же глаз, как у нас, с хрусталиком и со всем прочим? Этому мешает несколько довольно интересных причин. Прежде всего пчела слишком мала; если бы у нее был глаз, похожий на наш, но соответственно уменьшенный, то размер зрачка оказался бы порядка 30 мк, а поэтому дифракция была бы так велика, что пчела все равно не видела бы лучше. Слишком маленький глаз — это не очень хорошо. Потом, если сделать глаз большим, как голова пчелы, то он занял бы всю голову. Ведь ценность сложного глаза в том и состоит, что он практически не занимает места — просто тоненький слой на поверхности головы пчелы. Так что, прежде чем давать советы пчеле, не забывайте, что у нее есть свои собственные проблемы!
§ 5. Другие типы глаз
Кроме пчел, многие другие животные могут различать цвета. Рыбы, бабочки, птицы и пресмыкающиеся тоже могут различать цвета. А вот большинство млекопитающих, как полагают, не могут. Приматы, однако, различают. Птицы, несомненно, различают цвета, об этом говорит их окраска.
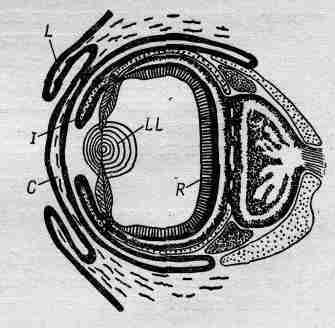
Фиг. 36.10. Глаз осьминога.
Какой был бы смысл самцам так блистательно ярко наряжаться, если бы самки не могли это видеть! Иначе говоря, привлекающее оперение, которым обладают птицы, и есть результат того, что самки способны различать цвета. Так что в следующий раз, когда вы увидите павлина и будете удивляться этой сверкающей выставке ярких красок, восхищаться утонченно подобранными цветами и замирать перед удивительным чувством эстетики птицы, не забудьте, что ваш восторг относится собственно к самке павлина, к ее наблюдательности и тонкому вкусу: только это ведь и породило столь удивительное зрелище!
Большинство беспозвоночных имеют либо недоразвитые, либо сложные глаза, а глаза всех позвоночных животных похожи на глаз человека. Однако есть одно исключение. Рассматривая высшие формы животных, мы обычно восклицаем: «Ну конечно, так и есть!», но если встать на менее предвзятую точку зрения и ограничиться только беспозвоночными, чтобы исключить нас самих, и спросить зоологов, какое из беспозвоночных животных они считают наиболее развитым, то большинство из них в один голос ответят — осьминог! Весьма интересно, что, помимо развитого мозга, его реакций и прочего, которые слишком хороши для беспозвоночного, осьминог имеет высокоразвитый глаз, совершенно непохожий на глаза кого-либо другого. Это не сложный глаз и не светочувствительное пятно, в нем есть и роговица, и веко, есть и радужка, и две полости, заполненные жидкостью, и хрусталик, и сетчатка (фиг. 36.10). В точности то же, что и у позвоночных! Это замечательный пример совпадения в эволюции, когда природа дважды пришла к одному и тому же решению проблемы, но с одним небольшим улучшением. Сетчатка у осьминога, как оказалось, представляет собой тоже часть мозга, и образовалась она при эмбриональном развитии, как у позвоночных животных, но имеется одно очень интересное и поразительное отличие: чувствительные к свету клетки расположены не позади слоев других клеток, как у нас, а непосредственно на внутренней поверхности глазного яблока, а клетки, занимающиеся вычислением,— позади них. Теперь мы по крайней мере видим, что в расположении клеток в нашем глазе глубокого смысла нет. В другой раз природе пришлось исправить свою ошибку! Самыми большими глазами наделен громадный спрут: диаметр их 38 см!
§ 6. Нервные механизмы зрения
Одной из основных тем этой главы является взаимосвязь и взаимоинформация отдельных частей глаза. Давайте рассмотрим сложный глаз краба-мечехвоста, над которым было проделано довольно много опытов.
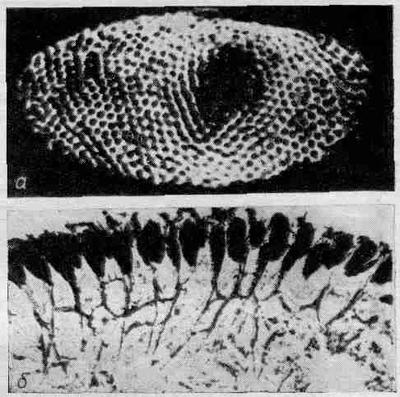
Фиг. 36.11. Сложный глаз краба-мечехвоста, а — общий вид; б — в разрезе.
Прежде всего нужно понять, какого сорта информация может передаваться по нервам. По нерву передается нечто вроде возмущения электрической природы, которое может быть легко зарегистрировано. Это некое волнообразное возмущение, которое бежит по нерву и вызывает на другом его конце какой-то эффект. Информацию переносит длинный отросток нервной клетки, называемый аксоном, и если один конец аксона возбужден, то по нему бежит «импульс». Далее, если по нерву уже проходит один импульс, то за ним не может немедленно последовать второй. Все импульсы имеют одну и ту же величину, так что, когда нерв сильно возбужден, это вовсе не означает, что по нему бежит больший импульс, а просто увеличивается, число импульсов в 1 сек. Величина же импульса определяется нервным волокном. Это важно усвоить, чтобы понять, что произойдет дальше.
На фиг. 36.11,а показан сложный глаз краба-мечехвоста; в нем всего лишь около тысячи омматидиев. Фиг. 36.11, б представляет собой поперечный разрез этой системы. Видны отдельные омматидии и нервные волокна, соединяющие их с мозгом. Но обратите внимание, что даже у этого краба имеются внутренние связи. Они, конечно, гораздо менее сложные, чем в глазе человека, но именно это-то и дает нам возможность изучить подобные связи на простом примере.
Давайте рассмотрим такой опыт: наложим на зрительный нерв нашего краба небольшие электроды и осветим только один омматидий; это легко можно сделать с помощью линз. Если мы в какой-то момент времени t0 включим свет и будем измерять возникающие электрические импульсы, то увидим, что после небольшой задержки последует быстрая серия разрядов, частота которых постепенно будет уменьшаться, пока не достигнет какой-то равномерности (фиг. 36.12,а). После выключения света разряды прекращаются. Интересно, что если усилитель остается связанным с тем же нервом, а мы направим свет на другой омматидий, то ничего не произойдет, сигналов не будет.
Проделаем теперь другой опыт: осветим первый омматидий и получим те же импульсы, но если теперь мы направим свет еще на другой соседний омматидий, то на короткое время импульсы прекращаются, после чего импульсы «побегут» снова, но с гораздо меньшей частотой (фиг. 36.12,б). Оказывается, что импульсы, возникающие во втором омматидий, затормаживают импульсы первого! Другими словами, хотя каждый нерв и несет информацию о своем омматидий, количество этой информации подавляется сигналами от другого омматидия. Например, когда более или менее равномерно освещен весь глаз, то сигнал, пришедший от любого отдельного омматидия, будет относительно слабым, ибо он подавлен множеством других сигналов. Торможение аддитивно, т. е. если мы осветим несколько соседних омматидиев, торможение будет очень сильным.
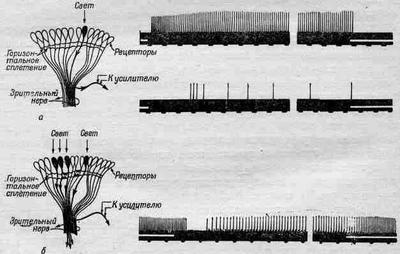
Фиг. 36.12. Ответная реакция на свет нервных волокон глаза краба-мечехвоста.
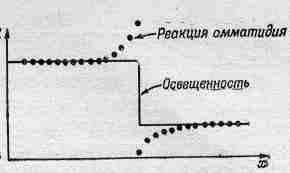
Фиг. 36.13. Ответная реакция омматидия краба-мечехвоста на резкий освещенный край.
Торможение оказывается большим, если омматидии расположены ближе, но если они достаточно удалены друг от друга, то торможение практически сводится к нулю. Таким образом, торможение аддитивно и зависит от расстояния. Это первый пример, когда информация от различных частей глаза перерабатывается в нем самом. Если подумать немного, то можно понять, что этот механизм предназначен для усиления контраста на краях объекта, ибо если часть предмета освещена, а часть нет, то омматидии, направленные на освещенную область, дают импульсы, которые затормаживаются всеми соседними омматидиями, видящими освещенную область, так что они относительно слабы. С другой стороны, омматидии, видящие границу световой области и дающие «белый» сигнал, хотя и подавлены своими соседями, но тех не столь уж много, ибо некоторые из них совсем темные (не видят света), в результате чего сигнал оказывается более сильным. В итоге получается кривая, подобная изображенной на фиг. 36.13. Краб как бы видит «усиленный» контур.
Однако тот факт, что существует такое «усиление» контура, известен уже давно. Это действительно замечательная вещь, которая неоднократно обсуждалась психологами. Чтобы изобразить предмет, нам достаточно нарисовать лишь его контур. Ведь мы привыкли видеть картины, на которых изображены одни только контуры! Но что же такое контур? Ведь это просто граница между темным и светлым или между одним цветом и другим. В самом деле, это вовсе не что-то определенное. Можно думать, что угодно, но никакой линии вокруг предмета нет! Нет, все это только наша выдумка. Теперь мы начинаем понимать, почему нам достаточно контура, чтобы представить себе весь предмет. По-видимому, наши глаза тоже работают, подобно глазам мечехвоста, разумеется, гораздо более сложно, но все же аналогично.
Наконец, я вкратце опишу более сложные опыты, очень красивые, и трудные, которые были проделаны над лягушкой. При выполнении их в зрительный нерв лягушки вводились миниатюрные искусно сделанные нитеобразные зонды и измерялись сигналы, идущие вдоль одного определенного аксона; точно так же как в случае с крабом-мечехвостом, было обнаружено, что информация зависит не просто от одной точки глаза, а является суммой информации, полученных от нескольких частей.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ПРОЧНОСТЬ ТВЕРДЫХ ТЕЛ И МЕХАНИЗМ ИХ ДЕФОРМАЦИИ
ПРОЧНОСТЬ ТВЕРДЫХ ТЕЛ И МЕХАНИЗМ ИХ ДЕФОРМАЦИИ Огромный вклад в эту чрезвычайно важную область физики внесли работы академика Абрама Федоровича Иоффе и его учеников.Большинство твердых тел имеет кристаллическую структуру. Долгое время в физике господствовали
24. Обманы зрения. Что длиннее?
24. Обманы зрения. Что длиннее? На приложенном здесь рисунке вы легко различите две главные линии: АВ и CD, по сторонам которых проведен ряд косых линий. Если вас спросят, какая линия длиннее, АВ или CD, – вы без колебания ответите:– Линия CD длиннее, чем АВ.Но вооружитесь
Все ли точки зрения равноценны?
Все ли точки зрения равноценны? Если бы наш интерес при наблюдении за движением тела в пространстве ограничивался изучением формы траектории (так называется кривая, по которой движется тело), то вопрос о выборе места наблюдения решался бы соображениями об удобстве и
2. Механизм каталитического взаимодействия. Виды катализаторов
2. Механизм каталитического взаимодействия. Виды катализаторов Различают гомогенный и гетерогенный катализ. При однородном (гомогенном) катализе катализатор и реагенты образуют однородную систему.Пример: представляют однородную газовую фазу. Границы раздела между
Механизм появления интерференции
Механизм появления интерференции Ученые всегда стремятся познать не только конечный и видимый результат, но и скрытые внутренние связи, протекающие в данном объекте, механизм, посредством которого реализуется результат. Это помогает им разобраться в наблюдаемом
МЕХАНИЗМ ХИГГСА
МЕХАНИЗМ ХИГГСА Ни один физик не сомневается в том, что на изученных нами до сих пор энергиях Стандартная модель работает. Результаты экспериментов согласуются с ее прогнозами с высокой точностью — лучше 1%.Однако Стандартная модель полагается на один ингредиент,
Механизм работы науки
Механизм работы науки Успех науки в изучении Вселенной складывается из наблюдений и выдвижения идей. Такого рода взаимообмен именуют научным методом (рис. 1.2). Рис. 1.2. Научный методВ ходе наблюдения то или иное явление воспринимается органами чувств при помощи приборов
Разные точки зрения на движение
Разные точки зрения на движение Чемодан лежит на полке вагона. В то же время он движется вместе с поездом. Дом стоит на Земле, но вместе с ней и движется. Про одно и то же тело можно сказать: движется прямолинейно, покоится, вращается. И все суждения будут верны, но с разных
Точка зрения звездного наблюдателя
Точка зрения звездного наблюдателя Мы решили изучать движение с точки зрения инерциальных систем. Не придется ли тогда отказаться от услуг земного наблюдателя? Ведь Земля вращается вокруг оси и вокруг Солнца, как доказал Коперник. Сейчас читателю, может быть, трудно
III. Движение с «неразумной» точки зрения
III. Движение с «неразумной» точки зрения
3.5. Динамика тел в Главном поясе. Механизм переноса вещества в область планет земной группы
3.5. Динамика тел в Главном поясе. Механизм переноса вещества в область планет земной группы Главный пояс астероидов — образование, имеющее сложную динамическую структуру. Эта структура в основном определяется силами, действующими на малые тела в этой области со стороны
Загадка нашего зрения
Загадка нашего зрения Вы уже знаете, что глаз устроен наподобие фотографической камеры и что на матовом стекле такой камеры все представляется в перевернутом виде.Вам известно также, почему мы видим предметы кругом нас не в перевернутом, а в настоящем их виде. С раннего
Обманы зрения на сцене
Обманы зрения на сцене Обманами зрения нередко пользуются фокусники, чтобы показывать на сцене свои необычайные «номера». Мне довелось раз присутствовать при борьбе фокусника с «привидением». Происходило это так:Вместе с другими я вошел в зал; мы сели и начали смотреть