Холиеэстераза и ацетилхолин
Согласно существующим представлениям, в основе механизма действия ФОС лежит избирательное торможение ими фермента ацетилхолинэстеразы, или просто холинэстеразы, которая катализирует гидролиз ацетилхолина-химического передатчика (медиатора) нервного возбуждения. Различают 2 типа холинэстеразы: истинную, содержащуюся преимущественно в тканях нервной системы, в скелетной мускулатуре, а также в эритроцитах, и ложную, содержащуюся главным образом в плазме крови, печени и некоторых других органах. Собственно ацетилхолинэстеразой является истинная, или специфическая, холинэстераза, так как только она гидролизует названный медиатор. И именно ее в дальнейшем мы будем обозначать термином «холинэстераза». Поскольку фермент и медиатор являются необходимыми химическими компонентами передачи нервных импульсов в синапсах — контактах между двумя нейронами или окончаниями нейрона и рецепторной клеткой, следует более подробно остановиться на их биохимической роли.
Ацетилхолин синтезируется из спирта холина и ацетилкоэнзима А[70] под влиянием фермента холинацетилазы в митохондриях нервных клеток и накапливается в окончаниях их отростков в виде пузырьков диаметром около 50 нм. Предполагается, что каждый такой пузырек содержит несколько тысяч молекул ацетилхолина. При этом в настоящее время принято различать ацетилхолин, готовый к секреции и расположенный в непосредственной близости от активной зоны, и ацетилхолин вне активной зоны, находящийся в состоянии равновесия с первым и не готовый к выделению в сипаптическую щель. Кроме того, имеется еще так называемый стабильный фонд ацетилхолина (до 15%), не освобождающегося даже в условиях блокады его синтеза.[71] Под воздействием нервного возбуждения и ионов Са2+ молекулы ацетилхолина переходят в синаптическую щель — пространство шириною 20–50 нм, отделяющее окончание нервного волокна (пресинаптическую мембрану) от иннервируемой клетки. На поверхности последней расположена постсинаптическая мембрана с холинорецепторами — специфическими белковыми структурами, способными взаимодействовать с ацетилхолином. Воздействие медиатора на холинорецептор приводит к деполяризации (снижению заряда), временному изменению проницаемости постсинаптической мембраны для положительно заряженных ионов Na+ и проникновению их внутрь клетки, что в свою очередь выравнивает потенциал напряжения на ее поверхности (оболочке).[72] Это дает начало новому импульсу в нейроне следующей ступени или вызывает деятельность клеток того или иного органа: мышцы, железы и др. (рис 5). Фармакологические исследования выявили существенную разницу в свойствах холинорецепторов различных синапсов. Рецепторы одной группы, проявляющие избирательную чувствительность к мускарину (яду гриба мухомора), названы мускариночувствительными, или М-холинорецепторами; они представлены главным образом в гладких мышцах глаз, бронхов, желудочно-кишечного тракта, в клетках потовых и пищеварительных желез, в сердечной мышце. Холинорецепторы второй группы возбуждаются малыми дозами никотина и поэтому названы никотино-чувствительными, или Н-холинорецепторами. К ним относятся рецепторы вегетативных ганглиев, скелетных мышц, мозгового слоя надпочечниковых желез, центральной нервной системы.
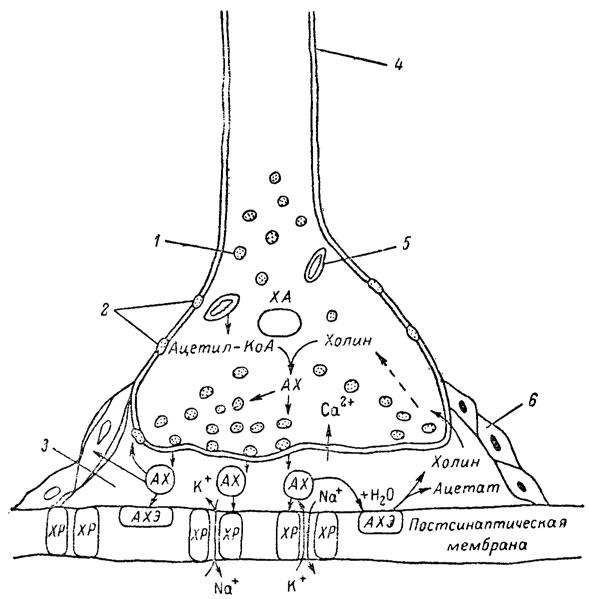
Рис. 5. Основные структурные элементы и схема функционирования холинергического синапса (Голиков, Фишзон-Рысс, 1978). 1 — синаптические пузырьки; 2 — пресинаптические центры связывания ацетилхолина; 3 — синаптическая щель; 4 — миелиновая оболочка; 5 — митохондрии; 6 — шванновские клетки; ХР — холинорецептор; АХ — ацетил-холин; АХЭ — ацетилхолинэстераза; ХА — холинацетилаза
Молекулы ацетилхолина, выполнившие свою медиаторную функцию, должны быть немедленно инактивированы, в противном случае будет нарушена дискретность в проведении нервного импульса и проявится избыточная функция холинорецептора. Именно это осуществляет холинэстераза, мгновенно гидролизующая ацетилхолин. Каталитическая активность холинэстеразы превышает почти все известные ферменты: по разным данным, время расщепления одной молекулы ацетилхолина составляет около одной миллисекунды, что соизмеримо со скоростью передачи нервного импульса. Осуществление столь мощного каталитического эффекта обеспечивается наличием в молекуле холинэстеразы определенных участков (активных центров), обладающих исключительно хорошо выраженной реакционной способностью по отношению к ацетилхолину.[73] Будучи простым белком (протеином), состоящим только из одних аминокислот, молекула холинэстеразы, как теперь выяснено, исходя из ее молекулярной массы, содержит от 30 до 50 таких активных центров.
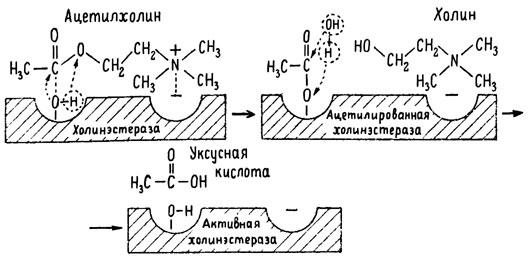
Рис. 6. Ферментативный гидролиз ацетилхолина. Объяснение в тексте
Как видно из рис. 6, участок поверхности холинэстеразы, непосредственно контактирующий с каждой молекулой медиатора, включает 2 центра, расположенных на расстоянии 0,4–0,5 мм: анионный, несущий отрицательный заряд, и эстеразный. Каждый из этих центров образован определенными группами атомов аминокислот, составляющих структуру фермента (гидроксилом, карбоксилом и др.). Ацетилхолин благодаря положительно заряженному атому азота (так называемой катионной головки) ориентируется за счет электростатических сил на поверхности холинэстеразы. При этом расстояние между атомом азота и кислотной группой медиатора соответствует расстоянию между активными центрами фермента. Анионный центр притягивает к себе катионную головку ацетилхолина и тем самым способствует сближению его эфирной группировки с эстеразным центром фермента. Затем рвется эфирная связь, ацетилхолин разделяется на 2 части: холиновую и уксусную, остаток уксусной кислоты присоединяется к эстеразному центру фермента и образуется так называемая ацетилирозанная холинэстераза. Этот крайне непрочный комплекс мгновенно подвергается спонтанному гидролизу, что освобождает фермент от остатка медиатора и приводит к образованию уксусной кислоты. С данного момента холииэстераза снова способна выполнять каталитическую функцию, а холин и уксусная кислота становятся исходными продуктами синтеза новых молекул ацетилхолина.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК